Charakteristickou črtou živých systémov, a teda aj bunky, sú rôzne druhy biologického pohybu: pohyb chromozómov v mitóze, premiestňovanie organel, prúdenie cytoplazmy, pohyb celých buniek. Tento pohyb sa uskutočňuje vďaka prítomnosti cytoskeletu. Cytoskelet je nepostrádateľná súčasť bunky, ktorá tvorí dynamickú trojrozmernú opornú sieť, pretkávajúcu celý cytosol. Cytoskelet zaisťuje bunke tvar, mechanickú stabilitu a pohyblivosť povrchových špecializácií, ako sú mikroklky, stereocílie a riasinky. Cytoskelet tvorí pohybovú aj opornú sústavu bunky. Základná cytoplazma je vyplnená zložkami cytoskeletu a poskytuje oporu pre bunkové organely. Existujú tri typy filament, ktoré tvoria cytoskelet, a to aktínové mikrofilamentá, intermediárne filamentá a mikrotubuly. Tieto sa skladajú z jednotlivých proteínov ovplyvňujúcich ich chemické aj fyzikálne vlastnosti. Jednotlivé filamentá môžu polymerizovať (predlžovať sa) alebo depolymerizovať (skracovať sa), čím umožňujú zmenu cytoskeletu v závislosti na potrebách bunky, napríklad pri bunkovom delení a pohybe bunky.
Hlavnou stavebnou súčasťou cytoskeletu je proteínové filamentum - polymérna dynamická štruktúra. Ďalšou súčasťou cytoskeletu sú doprovodné proteíny, ktoré regulujú stavbu a odbúravanie filament, prepájajú ich navzájom a pripájajú ich k iným štruktúram. Disociáciu a polymerizáciu jednotlivých filament ovplyvňujú doprovodné proteíny, ktoré sú pre každý typ filamenta špecifické.
Intermediárne filamenty: Pevnosť a odolnosť bunky
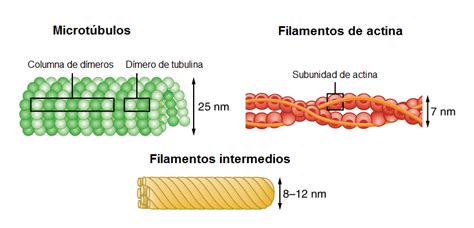
Intermediárne filamenty s priemerom 10-12 nm predstavujú rozsiahly systém vláknitých štruktúr bunky. Ide o mechanicky odolné vlákna. Sú tvorené dlhými molekulami, ktoré sú tvorené globulárnymi podjednotkami. Vyskytujú sa takmer vo všetkých bunkách, a to väčšinou v cytoplazme, niekedy aj v bunkovom jadre. Základnou podjednotkou je monomér intermediárneho filamenta, ktorý obsahuje tyčinkovitú oblasť a dva globulárne konce. Páry sa pomocou globulárnych koncov spájajú do dimérov. Dva diméry sa spoja paralelne s určitým posunom a vytvoria tetraméry, ktoré sa následne paralelne spájajú a vytvárajú stočenú štruktúru intermediárneho filamenta.
Intermediárne filamenty sú stabilné, pevné a tvoria oporu pre ostatné bunkové štruktúry. Považujú sa za zložku, ktorá má výhradne štrukturálnu funkciu (kostra bunky). Ide teda o nekontraktilné elementy. Bunke zabezpečujú mechanickú pevnosť, odolnosť na ťah a tlak. Vďaka svojej stabilite vyrovnávajú tlaky pôsobiace na bunku a dodávajú jej pevnosť. Zabraňujú deformácii buniek. Hrajú dôležitú úlohu v bunkovom delení. Zohrávajú úlohu aj pri transmembránovej signalizácii. Na rozdiel od dynamických mikrotubulov sú intermediárne filamenty zväčša permanentné štruktúry. V bunke slúžia ako pevné kotvy pre organely. Bunkové jadro zvyčajne visí v akejsi ochrannej „klietke“ vytvorenej práve z týchto vlákien.
Microtubules (Structure and Function)
Typy intermediárnych filamentov
Imunocytochemickými metódami bolo zistené, že intermediárne filamentá sú tvorené rôznymi typmi proteínov:
- Cytokeratínové intermediárne filamentá: Nachádzajú sa v epitelových bunkách, tvoria drobné zväzky - tzv. tonofibrily, ktoré sa viažu na desmozómy na laterálnych stenách. Delíme ich na kyslé, bázické aj neutrálne, z ktorých vždy dva typy tvoria polyméry.
- Vimentínové intermediárne vlákna: Nachádzajú sa v bunkách mezenchymálneho pôvodu, t.j. fixné bunky väziva, tukové bunky, osteocyty, chondrocyty, ďalej sa vyskytujú napr. v endotelových bunkách a bunkách obalu CNS.
- Neurofilamentá: Ich polyméry môžu byť tvorené tromi typmi proteínov (L, M a H).
V tomto mechanizme hrajú významnú úlohu aj proteíny asociované s jednotlivými typmi filament. U intermediárnych filament je to napríklad desmín. Mikrofilamentá sa spájajú ako navzájom, tak aj s intermediárnymi filamentami, ktoré sa nachádzajú v hlbšej vrstve.
Mikrofilamenty: Dynamické aktínové vlákna
Mikrofilamentá majú priemerne hrúbku 4 - 7 nm. Sú to najtenšie vlákna cytoskeletu, inak známe aj ako aktínové vlákna. Tvoria veľmi jemné špirálovité a kontraktilné (stiahnuteľné) reťazce. Aktínové mikrofilamentá s priemerom 5-7 nm sa nachádzajú vo všetkých eukaryotných bunkách, najväčší obsah aktínu má svalovina. Mikrofilamentá sa podieľajú zásadne na udržiavaní tvaru a vnútornej organizácie bunky.

Základnou jednotkou mikrofilamenta sú dve molekuly vláknitého F-aktínu, ktoré sa okolo seba obtáčajú a vytvárajú aktínové filamentum. Samotné vlákno F-aktínu sa skladá z polymerizovaných globulárnych podjednotiek G-aktínu. Základnou jednotkou je globulárny G-aktín (globulárny monomér) a fibrilárny F-aktín (polymerizujúci do asymetrickej dvojskrutkovice). Aktín sa vyskytuje v troch izoformách - α (kontraktilný aparát svaloviny), β a γ. Všetky globulárne molekuly aktínu smerujú vo vlákne rovnakým smerom pozdĺž osi vlákna. To vytvára v aktínových vláknach polaritu, a teda rozlíšiteľný plus (+) a mínus (-) koniec. V bunkách sa nachádza niekoľko typov aktínov, ktoré sa líšia aminokyselinami, ktoré ich tvoria. To spôsobuje rozdielnu funkčnú a štrukturálnu stabilitu.
Aktínové vlákna sa väčšinou vyskytujú vo zväzkoch, v sieťach alebo v rozvetvení. Aktínové filamentá, ktoré sa nachádzajú vo svalových bunkách, sú modifikované na tzv. myofilamentá a spojené pomocou tropomyozínu s hrubými myozínovými filamentami. Sú veľmi stabilné. Vo svalových bunkách sú na rozdiel od ostatných buniek lineárne a pravidelne usporiadané. V nesvalových bunkách sa vyskytuje ako aktín v polymerizovanej, tak v solubilnej forme a dochádza k neustálej polymerizácii a disociácii aktínových mikrofilament (polymerizáciu uľahčujú alebo blokujú profilíny 1 a 2).
Stupeň polymerizácie aktínu možno ovplyvniť zmenou hladiny iónov kalcia v cytoplazme. Polymerizácia aktínu prebieha tak, že aktínové monoméry v cytosole nesú ATP a po zapojení monoméru do rastúceho aktínového mikrofilamenta dôjde k hydrolýze na ADP. Molekuly ADP sú zachytené vo vnútri vlákna.
Microtubules (Structure and Function)
V nesvalových bunkách sú aktínové mikrofilamentá základom cytoskeletu. Tvoria difúznu sieť, ktorá je najčastejšie kondenzovaná pod bunkovou membránou v tzv. terminálnej sieti (membránový skelet). Terminálna sieť je naviazaná na bunkovú membránu pomocou proteínov spektrínu a dystrofínu. Samotné aktínové mikrofilamentá sú viazané k bunkovej membráne proteínmi α-aktínu, vinkulínu a tropomyozínu. Výsledný membránový skelet slúži hlavne na stabilizáciu bunkovej membrány a je naviazaný na zonula adherens. Mikrotrabekuly spájajú rozličné bunkové štruktúry navzájom aj s bunkovou membránou. Pokladajú sa za vlastnú kostru, oporu bunky. Aktínové mikrofilamentá sú nevyhnutné pre cytoplazmatický a bunkový pohyb. Sú súčasťou pseudopódií, lamellipódií a filopódií slúžiacich k migrácii bunky.
Myozín a aktín
Integrálnou zložkou mikrofilamentov je aj nesvalový myozín, ktorý sa od svalového odlišuje rozdielnym usporiadaním aminokyselín. Myozín predstavuje skupinu ATP-dependentných motorických proteínov, ktoré tvoria súčasť cytoskeletu všetkých živočíšnych a niektorých rastlinných eukaryotických buniek. Myozín II, ktorý je hojne zastúpený v ľudských bunkách, je tvorený 6 vláknami - 2 ťažkými a 4 ľahkými (2 regulačné a 2 esenciálne). Ťažké vlákna sa splietajú do dvojskrutkovice tvoriacej dlhý ocások. Globulárne hlavičky nasadajú na aktínové mikrofilamentá a kráčavým pohybom sa posúvajú smerom dopredu k plus (+) koncu filament. Výnimku tvoria myozín IV, VI, a IX pohybujúci sa v smere k mínus (-) koncu.
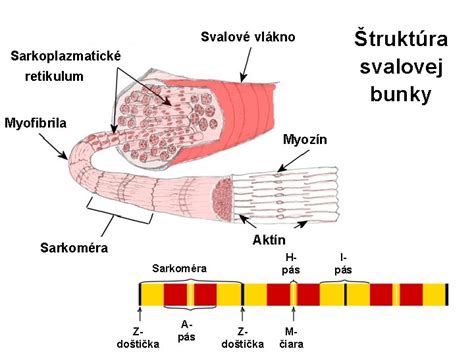
Dĺžka kroku je u myozínu jedného druhu konštantná, keďže sila pôsobí na krček pod stále rovnakým uhlom. Závisí ale na dĺžke krčku - čím dlhší krček je, tým dlhší je aj krok. Rozlišujeme 18 druhov myozínu značených rímskymi číslicami I - XVIII, ktoré tvoria „myozínovú superrodinu“ kódovanú asi 150 génmi (ľudský genóm obsahuje asi 40 génov kódujúcich myozín).
Mechanizmus kontrakcie v bunke je podobný, nie však identický so svalovou bunkou. Vyžaduje si prítomnosť energie z ATP a iónov Ca2+.
Mikrotubuly: Intracelulárny transport a bunkový pohyb
Mikrotubuly sú najhrubšie vlákna cytoskeletu (o priemere 25 nm), pozorovateľné aj vo svetelnom mikroskope. Sú to systém jemných tubulov s hrúbkou 20 - 40 nm a s lúmenom 10 - 20 nm. Mikrotubuly sú tubulárne (štruktúra "špirály"), pevné a nevětvící sa štruktúry, ktorých stena je tvorená 13 protofilamentami. Základným stavebným kameňom mikrotubulov je heterodimér tvorený α- a β-tubulínom. Základnou bielkovinou mikrotubulov je tubulín - heterodimér tvorený dvoma podjednotkami, ktoré sa líšia primárnou štruktúrou a označujú sa ako α, β.

Polymeráciou týchto dimérových molekúl (za účasti GTP) vznikajú špirálovito usporiadané protofilamentá, na ktorých rozlišujeme (+) a (−) koniec. Pri polymerizácii sa diméry organizujú takým spôsobom, že α-tubulín vždy susedí s β-tubulínom. Molekula tubulínu je polárna, má + a - koniec. Takéto chemické vlastnosti umožňujú vzájomné spájanie molekúl tubulínu a predlžovanie mikrotubulu v jednom smere (tzv. polárny rast). Polymerizáciou sa mikrotubuly predlžujú, depolymerizáciou skracujú. Zdrojom energie pre tento proces je guanozíntrifosfát (GTP, obdoba ATP). Nové podjednotky sa nachádzajú v depolymerizovanom stave v cytoplazme. Množstvo voľného tubulínu je v dynamickej rovnováhe s polymerizovaným tubulínom. Pri raste sa uplatňujú dva typy dimérov, jedny nesú GTP a druhé s naviazaným GDP. Mikrotubuly s naviazaným GTP pokračujú v raste tzv. GTP-čiapočka. Pokiaľ dôjde k hydrolýze GTP čiapočky na GDP, sú polyméry menej pevne viazané a dochádza k rozpadu a skracovaniu mikrotubulu. Rast mikrotubulov prebieha pridávaním heterodimérov na tzv. plus-konec. Pri polymerizácii hrá veľkú úlohu fosforylácia a defosforylácia doprovodných proteínov, ktoré sú k mikrotubulu pripojené. Aby nedošlo k depolymerácii mikrotubulu je nutné, aby boli oba konce chránené pred depolymeráciou.
Mikrotubuly sú buď nepravidelne usporiadané, alebo sú súčasťou bunkových štruktúr (deliace vretienko a centriola). Slúžia predovšetkým k intracelulárnemu transportu, ďalej umožňujú pohyb chromozómov a taktiež sa podieľajú na udržiavaní tvaru bunky a jeho zmenách. Zaisťujú hlavne tvar bunky (mechanická opora), ďalej sú nevyhnutné pre intracelulárny transport (napr. vezikuly, granulá a transport látok medzi endoplazmatickým retikulom a Golgiho komplexom) a sú súčasťou pohybového aparátu riasiniek a bičíkov. V priebehu mitózy vytvárajú mitotické vretienko a umožňujú tak pohyb chromozómov. Pri všetkých zmienených dejoch sú dôležité proteíny pripojené k mikrotubulám tzv. MAP (Microtubule Associated Proteins).
Microtubules (Structure and Function)
Centrozóm a bazálne telieska
V bunkách je proces novotvorby mikrotubulov (polymerizácia tubulínu) spojený s mikrotubulami organizujúcimi centrami (MTOC). Nové mikrotubuly vznikajú len v miestach MTOC: v živočíšnych bunkách je to centrozóm pre mitotické vretienko, bazálne telieska pre organizáciu mikrotubulov v riasinkách a bičíkoch. Centriol je komplexná štruktúra tvorená pravidelne usporiadanými mikrotubulami. Centriol pripomína tvarom dutý valec o dĺžke 0,3-0,5 µm. Stena centriolu je tvorená deviatimi tripletmi mikrotubulov, označených A, B a C. K centriolu nalieha tzv. oblasť pericentriolárneho materiálu, ktorý sa uplatňuje pri polymerizácii mikrotubulov (obsahuje prstence γ-tubulínu a proteíny potrebné k tvorbe mikrotubulu), ktoré sa radiálne šíria do okolitej cytoplazmy.
Bunka obsahuje jeden pár centriolov v blízkosti jadra a Golgiho komplexu (pokiaľ neprebieha bunkové delenie). Centrozóm je názov pre pár centriolov, ktoré sú na seba kolmé svojimi dlhými osami a pericentriolárny materiál. Centrozóm je niekedy nazývaný centrum organizujúce mikrotubuly (MTOC), pretože tu sú mikrotubuly ukotvené a vznikajú aj nové.
Motorické proteíny: Kinezíny a dyneíny
Pre intracelulárny transport sú mikrotubuly nezastupiteľné, pretože predstavujú vodiacu linku pre transportné častice, ktoré sa po nich pohybujú za pomoci tzv. motorických proteínov. Tieto molekuly majú dva póly, jeden pól obsahuje receptor naviazaný na transportné vezikuly a druhý s dvomi globulárnymi štruktúrami naviazanými na mikrotubul. Po naviazaní ATP dôjde na globulárnej časti k hydrolýze a uvoľnená energia slúži k zmene konfigurácie molekuly.
Motorické proteíny radíme do triedy tzv. molekulárnych motorov - molekúl, ktorých vlastnosťou bunka využíva k vnútrobunkovému transportu vezikul, granul a organel, prípadne k ďalším špecializovaným procesom (napr. svalový sťah, ciliárny aparát). Pre svoju funkciu vyžadujú chemickú energiu uvoľňovanú hydrolýzou najčastejšie ATP, menej často GTP, ktorú premieňajú v energiu mechanickú. Ako dráhu pre pohyb využívajú motorické proteíny cytoskeletárnu sieť - mikrofilamentá (aktín) alebo mikrotubuly.
Kinezíny
Kinezín je ďalší motorický proteín. Funkciou kinezínu je predovšetkým transport organel a vezikul. Molekula kinezínu sa líši podľa jednotlivých typov. Typický kinezín sa skladá z dvoch polymérov-ťažkých reťazcov, ktoré majú dva póly. Jeden pól obsahuje receptor s dvoma ľahkými reťazcami, ktorým sa pripája transportovaná partikula k motorickému proteínu. Po naviazaní ATP dôjde k hydrolýze a uvoľnená energia je použitá na zmenu konfigurácie kinezínu. Pri naviazaní jedného ATP sa proteín posunie o 8 nm. Kinezíny sa pohybujú smerom k (+) koncu mikrotubulu, hlavne po β-tubulínoch.
Dyneíny
Dyneín je motorický proteín, ktorý má za úlohu prenášať molekuly látok pohybom po koľajnici mikrotubulu smerom k jeho mínus (-) koncu. Tento spôsob prepravy sa nazýva retrográdny transport. Dyneíny pracujú ako tzv. molekulové motory. Dyneíny tvoria obrovské komplexy bielkovín, ktoré obsahujú niekoľko polypeptidových podjednotiek. Je dvojhlavá molekula s dvoma takmer identickými ťažkými reťazcami, ktoré tvoria domény hlavy a jedným ľahkým reťazcom, tvoriacim nôžku. Hlavy obsahujú aktivitu ATPázy a umožňujú tak pohyb pozdĺž mikrotubulu. Vyskytuje sa vo všetkých živočíšnych bunkách a jeho hlavnou funkciou je prenos vezikul a bunkových organel. Tým prispieva aj k umiestneniu jednotlivých organel v bunke.
tags: #intermediarne #filamenty #spajaju #denne #telieska